|
|
||||||
    |
|
|||||
EFECTOS DE LA DESAPARICIÓN DEL CULTIVO DE SECANO SOBRE EL MOCHUELO EUROPEO (Athene noctua)
Jose Antonio Martínez Climent * & Iñigo Zuberogoitia Arroyo**
C/ Juan de la Cierva 43, E-03560 El Campello, Alicante, España. E-mail: qvcocotiers@hotmail.com
**E.M. Icarus. Oficina Técnica. Apartado 106. 48940 Leioa. Bizkaia. E-mail: inigo.zuberogoitia@wanadoo.es
SUMMARY
In the period between 1993 and 2001 we have monitored changes in the number of territories
occupiedby Little Owls (Athene noctua) in four study-plots in San Juan (N of Alicante, Spain). The area was originally dominated by dry cultures (mostly Ceratonia siliqua). We report a 83,9 % decline in the number of occupied territories, and suggest a strong dependence between Little Owls and dry cultures, where the owls find nesting sites and suitable hunting habitat.
INTRODUCCION
La degradación o desaparición del hábitat de cría y caza es una de las principales causas últimas de la desaparición de varias especies de aves rapaces (THYOLLAY, 1993; MARTÍNEZ, et al 1999, entre otros). Por lo general, la mayoría de los estudios dedicados a evaluar estas relaciones se centran sobre especies y espacios protegidos o en vías de protección (eg. MARTÍNEZ, et al 1999), siendo escasa la atención que se presta a especies que se consideran abundantes aun a pesar de la ausencia de información precisa sobre el estado de sus poblaciones (SAUROLA, 1997).
El Mochuelo Europeo (Athene noctua) es una especie ampliamente distribuida en la Comunidad Valenciana, para la que no se ha descrito aspecto alguno de su ecología en dicha zona, y la escasez de estudios sobre esta especie es generalizada en toda la Península Ibérica (SEO/BirdLife, 1999). Como ocurre para otras aves rapaces, es frecuente emitir estimas académicas sobre su estatus y tendencias poblacionales que sobrevaloran en varios puntos porcentuales la realidad sobre su estatus a consecuencia del notorio comportamiento vocal del Mochuelo Europeo (SAUROLA, 1997).
El propósito de este trabajo es resumir las posibles relaciones entre la abundancia del Mochuelo Europeo y las variaciones en la superficie y composición de su hábitat preferente en una zona donde tradicionalmente se estima que la densidad de mochuelos es alta e invariable.
METODOS
Zona de estudio
La zona de estudio es la Huerta de San Juan (Alicante). Se sitúa al Norte de la ciudad de Alicante, entre el Río Montnegre al Oeste, Río Seco al Norte y Sierra Grossa al Sur, con el Mediterráneo al
Este. Este era, un paisaje dominado por los cultivos de olivo, almendro, algarrobo, pequeños viñedos y cereales de invierno. A pesar de tratarse de cultivos de secano, se le denomina “huerta” a causa de su eficiente sistema de recogida y aprovechamiento de las aguas, de origen islámico. Entre los cultivos se encontraban grandes haciendas familiares en las que era frecuente que anidaran los mochuelos (BUADES, com. pers.). Los años setenta suponen el inicio del desarrollo urbanístico incontrolado, y a mediados de los noventa se precipita la construcción de edificios, casas adosadas y carreteras que condicionan el declive ininterrupido del cultivo de secano.
Dentro de esta
zona, y a efectos de este estudio, se pueden distinguir cuatro parcelas
de cultivo de secano delimitadas por carreteras, pueblos y urbanizaciones.
La superficie ocupada por cada zona se puede apreciar en las Figura
1. Las distancias entre la Zona 1 (la más extensa) y las otras
tres oscila entre700m y 2,7 km.
Métodos de censo
Nuestro estudio se ha llevado a cabo entre 1993 y 2001. Con el fin de determinar con la mayor precisión posible el número y localización de territorios ocupados por los mochuelos europeos hemos seguido a ZUBEROGOITIA & CAMPOS (1998), MARTÍNEZ & ZUBEROGOITIA (2001) y ZUBEROGOITIA & MARTÍNEZ (2001). Durante los nueve años de trabajo de campo hemos llevado a cabo más de 1000 estaciones de escucha. Las escuchas se realizaron siempre en condiciones climáticas favorables para la escucha de cantos. Cada estación fue visitada tantas veces como fue necesario para confirmar la presencia o ausencia de un territorio ocupado. Cada día, las escuchas comenzaron al anochecer y se prolongaron entre 1 y 3 horas. Como método complementario, cada año hemos revisado todos los edificios y árboles susceptibles de albergar rapaces nocturnas.
Variaciones en la superficie y composición del hábitat
Las variaciones en la superficie de las 4 parcelas de estudio se cuantificaron utilizando planímetros sobre fotografías aéreas y cartografía 1:25.000. En la zona de estudio los mochuelos europeos crían preferentemente en orificios de algarrobos (Ceratonia siliqua) añejos. Por lo tanto, la descripción de las variaciones en la composición de las parcelas se han llevado a cabo contado cada año el número de pies de algarrobo (Ceratonia siliqua) añejo (con al menos un orificio susceptible de actuar como nido).
RESULTADOS
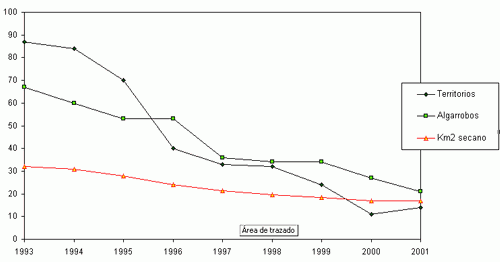
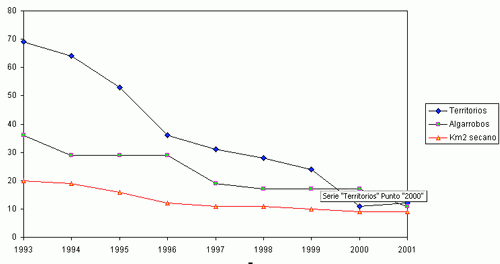
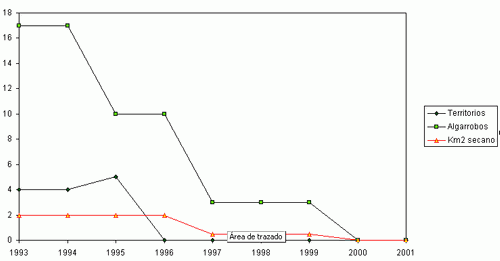
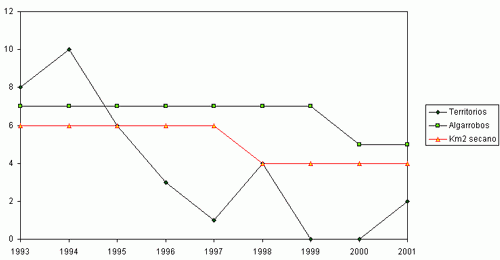
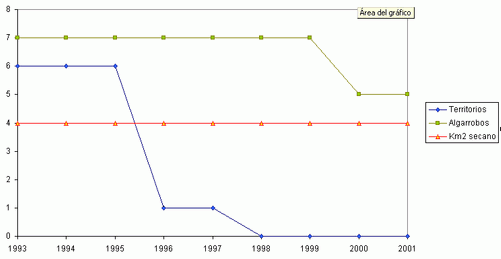
Durante el periodo comprendido entre 1993 y 2001 se ha constatado un declive del 83,9 % de la población original del Mochuelo Europeo en las estaciones de escucha situadas en Huerta de San Juan. La Figura 2 muestra la tendencia global del número de machos territorios detectados en los conteos primaverales realizados entre 1993 y 2001, así como la abundancia del número de pies de algarrobo añejo presentes en las parcelas de estudio y de la superficie de cultivo de secano en las mismas. Cada trío de puntos (en vertical) representa los valores para cada año. Es patente la fuerte correlación entre la abundancia de mochuelos, el cultivo de secano y los pies de algarrobo viejo. La Figura 1 (a, b, c y d) detalla las mismas relaciones para cada parcela de estudio. Para cada zona, y en su conjunto, las distintas correlaciones entre abundancia de mochuelos europeos, pies de algarrobo maduro y superficie de secano fueron significativas (pruebas r, P< 0,05).
DISCUSION
La Figura 2 muestra cómo la curva que marca la relación entre abundancia de algarrobos y mochuelos tiene una fuerte pendiente entre 84 y 33 pies, y esa pendiente no es tan evidente, para el mismo periodo de tiempo, en la relación entre superficie de secano y abundancia de mochuelos. Esto podría indicar la importancia de la disponibilidad de algarrobos añejos donde criar como factor limitante de la distribución del mochuelo. La situación se complica al analizar los resultados por parcelas de estudio. Nuestros resultados sugieren una pequeña estructura de metapoblaciones para el Mochuelo Europeo en la huerta de San Juan. Esta sugerencia se comenta siguiendo las recomendaciones y limitaciones propuestas por CRAIG (1986) respecto a “pequeñas estructuras metapoblacionales”. Estas estructuras locales son frecuentes en las poblaciones del Mochuelo Europeo en el resto de Europa (GÉNOT, 2001).
El fuerte declive en el número de pies de algarrobo viejo disponibles en la Zona 1 se ha visto
acompañado por la casi total desaparición de territorios ocupados en las otras zonas especialmente 3 y 4), en las que el declive en el número de algarrobos fue poco acusado. Las Zonas 2 a 4 se encuentran rodeadas por carreteras que soportan un gran flujo de vehículos, y en los arcenes de las mismas hemos encontrado numerosos mochuelos atropellados, pero únicamente en los primeros años de este estudio. Esto sugiere que la Zona 1, la de mayor superficie, podría actuar como “fuente”, como productora de mochuelos que luego habrán de dispersarse. Las otras zonas de estudio serían “sumideros”, pues la mortalidad producida por atropellos no es compensada por un flujo de mochuelos, ni provenientes de la Zona 1 ni de otros lugares. Es de esperar que la degradación o destrucción de la Zona 1 tenga severos efectos sobre la probabilidad de ocupación de territorios en la franja costera entre San Juan y Alicante a medio plazo. Como ocurre con otras especies de acusado comportamiento territorial (SAUROLA, 1997), el Mochuelo Europeo reafirma la posesión de su territorio frente a las nuevas cohortes durante los meses de septiembre-octubre, y posteriormente entre febrero y mayo (datos propios). En otoño se alcanza el máximo anual en el número de individuos (NEWTON, 1979; SAUROLA, 1997), lo que unido al considerable aumento de su detectabilidad suele inducir a sobrevalorar el tamaño real de la fracción reproductora de la población (SAUROLA, 1997). En Escandinavia, la desaparición de su hábitat preferido fuerza ocasionalmente al Búho real (Bubo bubo) a penetrar en cascos urbanos, siendo por lo tanto más dectectable. Como resultado, se produce una sobrevaloración del tamaño de sus poblaciones (Centro Finlandés de Anillamiento, com. pers.). Un razonamiento análogo podría explicar que la población de Mochuelos de la Huerta de San Juan se considere abundante y estable (lo que es contrario a los resultados de nuestros conteos sistemáticos), pues la desaparición de su hábitat fuerza probablemente a los mochuelos a visitar zonas aledañas, o incluso el interior, de los cascos urbanos. El Plan Parcial de Desarrollo Urbanístico de San Juan prevé la desaparición completa de los cultivos de secano remanentes en favor de urbanizaciones. Si bien el Mochuelo Europeo sigue siendo una especie abundante, cabe fomentar a partir de ahora la realización conteos sistemáticos en ecosistemas fragmentados de la Comunidad Valenciana.
AGRADECIMIENTOS
Estamos especialmente agradecidos al Sr. D. Isidro Buades, Cronista Oficial de la Villa de San Juan, por su ilustración sobre el paisaje tradicional de la huerta, y a Pertti Saurola (Centro Finlandés de Anillamiento) por revisar las versiones finesa e inglesa de esta trabajo.
REFERENCIAS
CRAIG, G. A. 1986. The Spotted Owl as framed by the timber industry. Western Timber Association and Artprint Press. Sacramento.
GÉNOT, J. C. 2001. État des connaissances sur la Chevêche `A-est de théna (Athene noctua) en bordure des Vosges du Nor (Nord-est de la France) de 1984 a 2000. Ciconia, 25 (2): 118. MARTÍNEZ, J. A., LÓPEZ, G., FALCÓ, F., CAMPO, A, & DE LA VEGA, A. 1999. Hábitat de caza y nidificación del Aguilucho Cenizo en el Parque Natural de la Mata-Torrevieja (Alicante, SE de España): efectos de la estructura de la vegetación y de la densidad de presas. Aredeola, 46: 205-212.
MARTÍNEZ, J. A. & ZUBEROGOITIA, I. 2001. The response of the Eagle Owl (Bubo bubo) to an outbreak of the rabbit haemorrhagic disease. Journal für Ornithologie, 142, 204-211.
NEWTON, I. 1979. Population Ecology of Raptors. Poyser, Carlton. London.
SAUROLA, 1997. Suomen pöllöt. Helsingin Uli. Helsinki.
SEO/BIRDLIFE, 1999. Seguimiento de Aves Nocturnas en España. Programa NOCTUA. Informe 1998. SEO/BirdLife, Madrid.
THYOLLAY, J. M. 1993. Response of a raptor community to shrinking area and degradation of tropical rain forest in the south western Ghâts (India). Ecography, 16, 97-110.
ZUBEROGOITIA, I. & CAMPOS, L. F. 1998. Censusing owls in large areas: a case study. Ardeola, 45, 47-53.
ZUBEROGOITIA, I. & MARTÍNEZ, J. A. 2001.The Little Owl in the Proyecto NOCTUA. Ciconia 25: 103-108.
Figura 1 (a, b, c, d). Variaciones anuales en la abundancia del Mochuelo Europeo en cada una de las parcelas de estudio en relación a la superficie de cultivo de secano y al número de pies de algarrobo añejo.
Figura 2. Variaciones anuales en la abundancia del Mochuelo Europeo en la Huerta de San Juan (Alicante) en relación con la superficie de cultivo de secano y al número de pies de algarrobo añejo. |
||||||
|
|
||||||